I just attended the openning of the 2011 symposium of Andy's Gonzalez's Quebec Centre for Biodiversity Science (http://qcbs.ca/). Michael Donoghue gave a cool keynote talk to start things off about the importance of studying and understanding both rapid and slow evolution if we are to confront the current biodiversity crisis. This blog will have more to say about the meeting in a future post but something prompted me to do a quick post starting a new "One from the Vault" series.
Blogs like this one tend to focus on new events and papers but it is fun to dig up old ones too. This new "One From the Vault" series will highlight older papers both from our lab and from other researchers. The current installment - Whither Adaptation? - was inspired by Andy's comment when we met after Michael's talk that he was amazed that Michael had mentioned the two of us in the same sentence. The surprise was that our views are often so different that it would seem unlikely that we would be lumped together under a common theme. But it did happen one other time ...
Whither adaptation?
by: Andrew P. Hendry and Andrew Gonzalez*
Abstract: The two authors of this paper have diametrically opposed views of the prevalence and strength of adaptation in nature. Hendry believes that adaptation can be seen almost everywhere and that evidence for it is overwhelming and ubiquitous. Gonzalez believes that adaptation is uncommon and that evidence for it is ambiguous at best. Neither author is certifiable to the knowledge of the other, leaving each to wonder where the other has his head buried. Extensive argument has revealed that each author thinks his own view is amply supported by both theory and empirical evidence. Further reflection has revealed that the differences in opinion may start with the different disciplines in which we work: evolutionary ecology for Hendry and community ecology for Gonzalez. In the present paper, we each present devastating evidence supporting our own position and thus refuting that of the other. We then identify the critical differences that led to such opposing views. We close by combining our two perspectives into a common framework based on the adaptive landscape, and thereby suggest means by which to assess the prevalence and strength of adaptation.
*Each author thinks the other contributed less
Hendry, A.P., and A. Gonzalez. 2008. Whither adaptation? Biology and Philosophy 23:673-699.
http://www.springerlink.com/content/u741773914w3k2w7/

Academic musing by Hendry, Bolnick, Gotanda, and awesome guests. Opinions and statements expressed on this blog are the views of the author(s) and do not necessarily represent endorsement by the blog administrators.
Thursday, December 8, 2011
Monday, December 5, 2011
Sunday, December 4, 2011
Non-parallel evolution in Storrs and stickleback
Independent populations that colonize similar environments and then evolve similar traits provide strong evidence for a deterministic response to natural selection. The resulting pattern has been variously called “parallel” or “convergent” depending on similarity among the starting populations and on one’s preferred definition.*
As parallel evolution provides some of the most convincing evidence for the power of natural selection, finding it has become a rite of passage for evolutionary biologists working in any given natural system, and it is a common stamp of approval for authors publishing on adaptation. Over the years, I have probably seen more than a hundred papers that examine phenotypes from multiple populations in two or more environments. Some statistical model is applied to the data and the main effect of “habitat type” is examined with great expectation. When this effect is significant, the paper trumpets its success in documenting the power of natural selection in shaping the diversity of life.
What often strikes me about such papers, however, is that the population means of the different habitat types often overlap considerably. That is, some populations in one habitat type have larger traits values and some have smaller trait values than populations in the other habitat type. This pattern implies to me that the coarse classification of habitat type is, in reality, not very predictive of phenotype and that many other factors must also be involved. Sadly, investigators rarely embrace and explore – or even acknowledge – this unexplained variation. We recently published a paper* that does so, and I was prompted to make a blog entry about it as a result of my seminar visit last week to the University of Connecticut.
Parallel/convergent evolution was a common theme in discussions with many UConn folks. Some labs studied situations where different organisms occupy similar environments and so would be expected to evolve similar traits (Schlutz lab: adaptation to the osmoregulatory challenges of fresh versus salt water by stickleback and alewives). Other labs studied situations where similar body forms evolved independently and so one would expect similar selective pressures to have been the cause (Jockush lab: large and small bodied skinks of different lineages in California). In South Africa, the plant genus Pelargonium is highly variable in leaf shape and growth form (Schlichting and Holsinger labs). So variable, in fact, that I (as a non-plant person) would never have imagined that they would all be in the same genus. Presumably all of these different shapes and forms in Pelargonium are the result of adaptation to different environments, and yet species with very different growth forms can be found side-by-side in the same environment. (OK, I only saw them in a greenhouse but I am told this is true in nature.) If adaptation is the cause of this group’s radiation, then why are two plant types of very different form found in the same environment? This dramatic non-parallelism is just the sort of thing I am talking about.
To investigate non-parallelism, Renaud Kaeuffer, Dan Bolnick, Katie Peichel, and I quantified ecological (diet), morphological (body shape, trophic traits, armor traits), and genetic (neutral and QTL-linked microsatellites) variation in each of six independent lake and stream population pairs of threespine stickleback*. Some traits were remarkably parallel. For example, all lake populations had stickleback with shallower average body depths than did all stream populations (no overlap in the population means). Other traits, however, were decidedly non-parallel. For example, the main effect of habitat type (lake or stream) explained essentially none of the variation in armor traits. So what is the cause of this variation in parallelism? By relating divergence in morphological traits to the degree of ecological divergence, we were able to show that deviations from non-parallelism are mostly likely the result of variation in selection. That is, variation in parallelism is caused by selective factors that do not map so cleanly onto the coarse habitat type classifications that we (and possibly other investigators) typically use. We are not the first group to find this, of course, but we certainly have fun embracing it.
The pressing question to my mind is just how common and strong are these effects? How similar will be the physiological adaptation of alewives and stickleback? To what extent is the variation in skink body shape the result of adaptation to particular environments – as opposed to some other effect? Is the variation in growth form in Pelargonium the result of variation in microhabitats with a general location – or is it the result of some other historical contingency? To what extent does variation in gene flow cause deviations from parallel evolution. We previously found this last effect to be strong in lake and stream stickleback, and it also seems critical in Mark Urban’s salamanders.
Just how parallel is parallel evolution – and why?
As parallel evolution provides some of the most convincing evidence for the power of natural selection, finding it has become a rite of passage for evolutionary biologists working in any given natural system, and it is a common stamp of approval for authors publishing on adaptation. Over the years, I have probably seen more than a hundred papers that examine phenotypes from multiple populations in two or more environments. Some statistical model is applied to the data and the main effect of “habitat type” is examined with great expectation. When this effect is significant, the paper trumpets its success in documenting the power of natural selection in shaping the diversity of life.
What often strikes me about such papers, however, is that the population means of the different habitat types often overlap considerably. That is, some populations in one habitat type have larger traits values and some have smaller trait values than populations in the other habitat type. This pattern implies to me that the coarse classification of habitat type is, in reality, not very predictive of phenotype and that many other factors must also be involved. Sadly, investigators rarely embrace and explore – or even acknowledge – this unexplained variation. We recently published a paper* that does so, and I was prompted to make a blog entry about it as a result of my seminar visit last week to the University of Connecticut.
Parallel/convergent evolution was a common theme in discussions with many UConn folks. Some labs studied situations where different organisms occupy similar environments and so would be expected to evolve similar traits (Schlutz lab: adaptation to the osmoregulatory challenges of fresh versus salt water by stickleback and alewives). Other labs studied situations where similar body forms evolved independently and so one would expect similar selective pressures to have been the cause (Jockush lab: large and small bodied skinks of different lineages in California). In South Africa, the plant genus Pelargonium is highly variable in leaf shape and growth form (Schlichting and Holsinger labs). So variable, in fact, that I (as a non-plant person) would never have imagined that they would all be in the same genus. Presumably all of these different shapes and forms in Pelargonium are the result of adaptation to different environments, and yet species with very different growth forms can be found side-by-side in the same environment. (OK, I only saw them in a greenhouse but I am told this is true in nature.) If adaptation is the cause of this group’s radiation, then why are two plant types of very different form found in the same environment? This dramatic non-parallelism is just the sort of thing I am talking about.
To investigate non-parallelism, Renaud Kaeuffer, Dan Bolnick, Katie Peichel, and I quantified ecological (diet), morphological (body shape, trophic traits, armor traits), and genetic (neutral and QTL-linked microsatellites) variation in each of six independent lake and stream population pairs of threespine stickleback*. Some traits were remarkably parallel. For example, all lake populations had stickleback with shallower average body depths than did all stream populations (no overlap in the population means). Other traits, however, were decidedly non-parallel. For example, the main effect of habitat type (lake or stream) explained essentially none of the variation in armor traits. So what is the cause of this variation in parallelism? By relating divergence in morphological traits to the degree of ecological divergence, we were able to show that deviations from non-parallelism are mostly likely the result of variation in selection. That is, variation in parallelism is caused by selective factors that do not map so cleanly onto the coarse habitat type classifications that we (and possibly other investigators) typically use. We are not the first group to find this, of course, but we certainly have fun embracing it.
The pressing question to my mind is just how common and strong are these effects? How similar will be the physiological adaptation of alewives and stickleback? To what extent is the variation in skink body shape the result of adaptation to particular environments – as opposed to some other effect? Is the variation in growth form in Pelargonium the result of variation in microhabitats with a general location – or is it the result of some other historical contingency? To what extent does variation in gene flow cause deviations from parallel evolution. We previously found this last effect to be strong in lake and stream stickleback, and it also seems critical in Mark Urban’s salamanders.
Just how parallel is parallel evolution – and why?
Thursday, November 17, 2011
Why natural selection will never cure the common cold!
In my last blog entry, I described a trip to Uppsala, Sweden, and the Beaver Snaps I drank there. I learned at the time that Beaver Snaps are often a northern Swedish speciality. This last week, I returned to Sweden – this time much closer to the land of Beaver Snaps. I was in Umea for their annual KBC days, an event where researchers from several departments at the university gather together to interact and share their research findings. My job was to give a plenary talk of general interest – and I spoke on Humans, Evolution, and the Future of Biodiversity. This was simply a grandiose title for what amounted to an advertisement for the importance of considering evolution when evaluating how human activities will shape future biodiversity. I also gave a research talk about ecological speciation.
The cool thing about trips like this is that you talk to all sorts of crazy people who have all sorts of crazy ideas. These ideas are sometimes useful and inspiring and sometimes not, but they are certainly always interesting. And sometimes these crazy ideas are what I remember most, such as in my “What did I learn today” entry from Belgium last winter. On this most recent trip, I spent some hours arguing about behavioural syndromes with my host Tomas Brodin – I remain to be convinced! And Lennart Persson espoused his paradoxical idea – they even have theory and data! – that increasing mortality on a given age class will increase the number and biomass of individuals in that age class. Already published in Science and PNAS, this theory is coming soon to a bookstore near you.
Now that stuff was bizarre enough but perhaps the most surprising discussion started with a question from someone who had seen my general talk. He wanted to know why natural selection has not eliminated our susceptibility to colds – after all they do make us damn miserable and we seem to get them all the time, particularly those of us with kids (more about this later). Now the usual answer to this question might be that colds are caused by micro-organisms that have very short generation times that allow them to adapt to us better than we can adapt to them. But I figure the more interesting answer is that colds don’t currently reduce our fitness – so selection won’t act to eliminate them. I certainly had a raging cold at the time and so the question stuck with me. I began to wonder if colds had actually reduced my reproductive success. I have two kids now but, without colds, would I have three – or four?
So clearly we need a study on this. Perhaps we should ask people whether or not they conceived their children when they had a cold. I suppose that would be hard to remember (hopefully so anyway) and so maybe we could instead test whether people that get more colds have fewer children. Better yet, we could experimentally induce colds in some bars and not others (that should be easy – we just infiltrate the dish washing staff) and then monitor mating and reproductive rates of the patrons (probably a bit harder). And, to figure out if sexual selection was the reason, we could take a bunch of people with colds, give some decongestants and not others, and send them into bars with instructions to ... well you get it. And why not take this train of thought to its logically absurd conclusion. I noted earlier that people with kids get more colds than do people without. Maybe this is a case of selection through sibling conflict. It clearly benefits the kids you already have if you don’t have more kids – because the available resources are divided to a lesser extent. So maybe, just maybe, colds are adaptive in children because they enable them to infect their parents thus (further) reducing the chance the parents will have more kids. Sick kids really are the birth control we always thought they were!

My first daughter, Aspen, trying hard to make sure I don't have a second daughter. It didn't work - perhaps because she couldn't have a cold all the time.
The cool thing about trips like this is that you talk to all sorts of crazy people who have all sorts of crazy ideas. These ideas are sometimes useful and inspiring and sometimes not, but they are certainly always interesting. And sometimes these crazy ideas are what I remember most, such as in my “What did I learn today” entry from Belgium last winter. On this most recent trip, I spent some hours arguing about behavioural syndromes with my host Tomas Brodin – I remain to be convinced! And Lennart Persson espoused his paradoxical idea – they even have theory and data! – that increasing mortality on a given age class will increase the number and biomass of individuals in that age class. Already published in Science and PNAS, this theory is coming soon to a bookstore near you.
Now that stuff was bizarre enough but perhaps the most surprising discussion started with a question from someone who had seen my general talk. He wanted to know why natural selection has not eliminated our susceptibility to colds – after all they do make us damn miserable and we seem to get them all the time, particularly those of us with kids (more about this later). Now the usual answer to this question might be that colds are caused by micro-organisms that have very short generation times that allow them to adapt to us better than we can adapt to them. But I figure the more interesting answer is that colds don’t currently reduce our fitness – so selection won’t act to eliminate them. I certainly had a raging cold at the time and so the question stuck with me. I began to wonder if colds had actually reduced my reproductive success. I have two kids now but, without colds, would I have three – or four?
So clearly we need a study on this. Perhaps we should ask people whether or not they conceived their children when they had a cold. I suppose that would be hard to remember (hopefully so anyway) and so maybe we could instead test whether people that get more colds have fewer children. Better yet, we could experimentally induce colds in some bars and not others (that should be easy – we just infiltrate the dish washing staff) and then monitor mating and reproductive rates of the patrons (probably a bit harder). And, to figure out if sexual selection was the reason, we could take a bunch of people with colds, give some decongestants and not others, and send them into bars with instructions to ... well you get it. And why not take this train of thought to its logically absurd conclusion. I noted earlier that people with kids get more colds than do people without. Maybe this is a case of selection through sibling conflict. It clearly benefits the kids you already have if you don’t have more kids – because the available resources are divided to a lesser extent. So maybe, just maybe, colds are adaptive in children because they enable them to infect their parents thus (further) reducing the chance the parents will have more kids. Sick kids really are the birth control we always thought they were!
My first daughter, Aspen, trying hard to make sure I don't have a second daughter. It didn't work - perhaps because she couldn't have a cold all the time.
Tuesday, November 15, 2011
Joining the Carnival!
There's a "blog carnival" called the Carnival of Evolution that hosts a monthly roundup of the best evolution-related blog posts from across the web. Having just discovered this, our blog will be joining the fun, starting next month. For now, you can read the entries for the current carnival, #41, at the mermaid's tale. It's a great way to discover evolution blogs you didn't know existed, and a great way to see editor-chosen "best of" picks for the whole month.
Enjoy!
Enjoy!
Friday, November 11, 2011
Fates Beyond Traits
Many entries on this blog focus on the impacts humans are having on evolutionary processes. The recent description of the impact of human activity on Darwin's finch evolution is one such example. But do human impacts on eco-evolutionary dynamics stop there - on the traits themselves? Or can human-induced trait changes have knock-on effects on ecological processes?
In a recent paper, my coauthors and I explore the evidence that human-induced trait changes in wild populations are impacting ecological processes. We call these ecological consequences "fates beyond traits." We conclude that human-induced trait changes may be impacting ecological processes on a global scale, but that more work is needed to understand the nature of these effects.
Please check out some examples of fates beyond traits on my lab website.
Palkovacs, E.P., M.T. Kinnison, C. Correa, C.M. Dalton & A.P. Hendry. 2011. Fates beyond traits: ecological consequences of human-induced trait change. Evolutionary Applications. Online early. doi:10.1111/j.1752-4571.2011.00212.x
Monday, October 31, 2011
Magic traits, muggle traits and squib traits
Speciation is a tricky business. Let's consider ecological speciation [1], a broad term for speciation that is the result of adaptation to different environments. The idea here is that 1) different environments generate divergent selective pressures on organisms, 2) this divergent selection causes divergent adaptations to the different environments, and 3) this divergent adaptation then leads to reproductive isolation, by various mechanisms. (The different "environments" might be in different places, at least partially geographically isolated from each other, or they might actually be different habitats or niches in a single place, such as different plants in a single meadow that plant-eating insects might adapt to eating; I'm going to gloss over this "allopatry" versus "sympatry" distinction here since it isn't particularly important to the points I'm going to discuss.)
Here's a specific empirical example: one of Darwin's finches, the Medium Ground Finch (Geospiza fortis) of the Galápagos (Fig. 1; discussed more in the previous post on this blog). We'll consider these birds as an example of "speciation in progress," but of course this means that it might not be an example of speciation at all – the early stages of divergence observed in this system may never "run to completion" and produce two fully distinct species. But because speciation is a process, it's best understood by looking at it as it's happening; by the time it's complete, much of the evidence regarding exactly how the process unfolded has vanished.

To make a long story short, and glossing over many important details (see that previous blog entry and [1-4] for further discussion and cites), in the Galápagos there are small seeds and large seeds. The finches eat the seeds, and their beak size matters for this purpose because a large beak can generate more bite force than a small beak. So small-beaked birds are better suited to cracking small seeds, and large-beaked birds are better suited to cracking large seeds. The environment, by offering up these two seed types, is therefore applying divergent selection to the finches, and the finches, obediently following Darwin, have diverged adaptively: there are small-beaked morphs and large-beaked morphs, and the phenotypic distribution is bimodal, meaning there are relatively few medium-beaked morphs.
So that's great. Divergent selection, divergent adaptation – speciation? Not so fast. The problem is that sexual species, like these finches, mate, and when they mate, their offspring inherit chromosomes from both parents, which leads (in the typical case) to intermediate phenotypes. In other words, when a big-beaked finch mates with a small-beaked finch, their hybrid offspring will tend to be medium-beaked. So sexual reproduction pulls a population back together, opposing the diversifying force of divergent selection. This is the reason why the most commonly used definition of "species," the "biological species concept," defines "species" as being (mostly) reproductively isolated; if you're not reproductively isolated, then there's always the possibility that your incipient diversity will collapse back into a single phenotype due to mating.
How can this problem be overcome? This is the third and final step of ecological speciation, as I set it out at the beginning of the post: divergent adaptation must somehow lead to reproductive isolation. The finches seem to manage this: in some populations, there are relatively few medium-beaked morphs. There are lots of ways in which this can be achieved, and that's really what I want to talk about in this blog entry. Back in 2004, Sergey Gavrilets coined the term "magic trait" in a now-famous passage:
In that passage, Gavrilets gave a solution as well: establishing that correlation is much easier if the loci under divergent selection and the loci that affect non-random mating are one and the same. Then divergent adaptation leads inevitably to non-random mating that allows the divergence to be maintained. It's like magic! (Fig. 2)

But there are several things about this definition that are hard to pin down, and so confusion reigned o'er the land until Servedio et al. bravely stepped into the fray [3]. In their much-needed paper, they clarify this definition in several important respects. First of all, there's the confusion over whether a magic trait is a phenotypic trait, as commonly imagined with reference to traits like beak size (and there are many other examples; see [3]), or whether it's actually a "magic gene" that governs such a trait, or perhaps even a set of "magic genes" (note Gavrilets' use of the word "loci," plural). I'll let you read [3] to see how they handled this hot potato. Second of all, there was a thorny problem regarding exactly which sorts of traits would be considered "magic" and which would not. Servedio et al. resolved this by defining two types of magic traits, "automatic" and "classic," which I'll explain with reference, again, to G. fortis.
Consider a large-beaked finch and a small-beaked finch that mate and have a medium-beaked baby finch (aww, isn't it cute?): that baby finch is the problem, and it must be eliminated. How to eliminate it? (I am speaking loosely for rhetorical purposes; I mean that it must be eliminated if divergence is to proceed, but of course G. fortis is not planning all this, nor is evolution guided somehow towards a goal of speciation.) One mechanism is via selection against that hybrid. If there are no medium-sized seeds in the system, then our medium-beaked bird will probably be outcompeted, will have low fitness, and will be relatively likely to die before reproducing. Through this mechanism, beak size is an "automatic magic trait"; reproductive isolation occurs, via selection against hybrids, as an automatic consequence of beak size being under divergent selection. There are other mechanisms by which magic traits can be automatic, involving spatial or temporal isolation, habitat choice, and some kinds of pollinator-related dynamics; see [3] for more details.
The other way that this undesirable baby bird can be eliminated (again, speaking loosely) is by not having it in the first place, through mate choice. If small-beaked finches prefer to mate with other small-beaked finches, and large-beaked finches prefer to mate with other large-beaked finches, then fewer hybrids will be produced (perhaps even none). And in fact, G. fortis does seem to mate assortatively, small with small and large with large, both because of a visual preference based on beak size, and because of imprinting on parental song, which, because beak size affects song, leads to a preference for mates with a beak size similar to one's own. This (both of these mechanisms of assortative mating based on beak size) makes beak size also a "classic" magic trait, by the definition of Servedio et al. The key distinction is that the non-random mating here is not inevitable or automatic; even with beak size under divergent selection, there is no a priori reason to think that that will lead to assortative mating. Instead, a separate genetic mechanism of some kind must be in place that establishes the rules that make it work, a mechanism that produces behaviors such as "I prefer to mate with males that have large beaks" or "I prefer to mate with males that sing a song like the one I heard my father sing when I was a baby." (I am here glossing over a further complication, between one-allele and two-allele mechanisms; see [3] and Felsenstein 1981). Once those rules, saying in one way or another "use beak size as a male mating cue," are set up (and they may have been set up in an ancestral species, long before G. fortis evolved), beak size acts as a magic trait; but since the use of beak size as a mating cue depends on that separate, external mechanism, it can be broken down, so it is not "automatic," it is "classic."
So far so good. Now our finches are 1) under divergent selection, 2) they exhibit adaptive divergence in response, and 3) they can preserve that adaptive divergence, to at least some extent, because the trait that is diverging, beak size, also "magically" produces assortative mating through both an automatic mechanism (selection against medium-beaked hybrids) and two classic mechanisms (female preference for particular beak sizes, and female preference for songs similar to the parental song they imprinted on). So... speciation? Again, not so fast.
The problem is that there are still various ambiguities and problems surrounding the "magic trait" definition, and so I and four co-authors wrote a letter to TREE [4] in response to Servedio et al. (and Servedio et al. replied [5]). Let's look at these difficulties, continuing with G. fortis as our example (see [4] for cites in support).
First of all, it's hard to decide whether beak size in G. fortis is truly "magic" or not, even by the clarified definition of Servedio et al. In G. fortis, divergent selection is only present in some years (and only at some sites); in other years (or at other sites), particularly when seeds are plentiful and so competition is low, medium-beaked individuals seem to do fine. So the black-and-white requirement of a "trait under divergent selection" is fulfilled only some of the time; is that good enough or not? In our letter, we propose that the "magicness" of magic traits is really the result of a trait-environment interaction. Beak size is not a "magic trait" unless it is placed in an environment that subjects it to divergent selection. In other environments, it is a "muggle trait," a trait which influences non-random mating but which is not presently under divergent selection; if a muggle trait is subjected to divergent selection, it becomes a magic trait. "Muggle trait" is a term coined by Eva Kisdi following the Harry Potter terminology in which "muggles" are people that are non-magical (Fig. 3). Similarly, such "magic environments" are only magic for organisms with a beak size trait that is used as a mating cue. So there will be, then, a spatiotemporal mosaic of "magicness" in those spots where the trait and the environment come together. It doesn't really make sense to talk about a "magic trait" except with reference to the "magic environment" it is in.

Whether beak size in G. fortis is magic is unknown for other reasons as well. The "automatic" part of the non-random mating, since it depends upon selection against hybrids, occurs only for some years/sites, as just explained. The "classic" aspects of non-random mating in G. fortis depend upon how consistently, through time and across space, beak size is used as a mating cue, and that has not been studied well enough to draw firm conclusions, to my knowledge; it may be, for example, that beak size is only important for mate choice in lean years. The requirement of pleiotropy is also on slightly shaky ground, although it's better established than other aspects. Beak size certainly seems to be the trait under divergent selection, and seems also to be the trait used as a mating cue, and so pleiotropy seems guaranteed (since beak size and beak size are the very same phenotypic trait). But the hypothesis that beak size is the true trait subject to both divergent selection and assortative mate choice (rather than a correlated trait) can only really be shown conclusively with manipulative experiments (altering beak size somehow and observing the effect on fitness and mate choice) that are impossible to conduct since the Darwin's finches are protected species; so pleiotropy is not conclusively proved.
So we can't actually say whether beak size is magic or not; indeed, we can't even say with certainty whether any of the three definitional requirements are satisfied, despite this being one of the most extensively studied species on the planet. But there's a problem in the other direction as well: it might be a little too easy to claim that beak size is magic. You see, the definition of "magic trait" says nothing about the magnitude of any of its requirements. Even the tiniest amount of "divergent selection" greater than zero suffices. Even the tiniest tendency towards "non-random mating" above panmixia suffices. And even the tiniest amount of pleiotropy suffices, too – one could imagine an organism for which nose length is under divergent selection, toenail growth rate is used as a mating cue, and these two traits are largely governed by independent loci, but there is one locus, perhaps a regulatory gene of some sort, that has a tiny pleiotropic effect on both. That regulatory gene would then be a "magic trait" despite its very small pleiotropic contribution to the two phenotypic traits in question. So the definition draws a line, but at a place that isn't really related to the function of the thing being defined.
There are lots of cases where we work with such definitions. The car up on cinderblocks in your neighbor's yard had its wheels taken off a couple years back, and its engine is probably rusted firmly together; but it's still a "car," even though it isn't usable as transportation (Fig. 4). On the other hand, we often choose not to use such definitions. The "biological species concept" (BSC) definition of "species," set out by Ernst Mayr, originally required complete reproductive isolation between species, because otherwise two putative species might collapse back into a single species, in a case of the random mating problem we've been discussing, writ large. But in later writing Mayr (and the rest of biology) backed off from that requirement, as it became clear that many species in nature actually do hybridize with other species to a limited extent, and thus the absolute definition would not draw a line in a useful place. Rather than trying to draw a new, arbitrary line regarding just how much hybridization is allowable, the working definition of the biological species concept now essentially allows a vague "you know it when you see it" amount of hybridization. Despite this vagueness, it has proved more useful than the old, absolute definition. (Similarly, if you asked your neighbor "Hey, can I borrow your car?" and he gave you the keys to the Chevy on blocks, you would probably be unhappy – and if this happened frequently, you might start considering a different definition for the word "car"!)

Another reason the original BSC definition of "species" was loosened was that it wasn't testable. How do you prove that two species positively, absolutely, never, ever, interbreed? That's unprovable, and so the definition, used strictly, was useless. Similarly, how do you prove that a given trait 1) is not subject to even the tiniest smidge of divergent selection, 2) absolutely never affects mate choice, and 3) is not governed, even to the tiniest degree, by pleiotropy at any locus? You could never prove such a proposition, for any trait, and so for all we know, every single trait in every single (sexual) organism is magic. In fact, that's not even an unreasonable thing to assert. I think it might even be true. So maybe that's a problem.
This starts to get at the heart of the problem: effect size. The effect size of a magic trait is "how much the trait contributed to the evolution of increased reproductive isolation" [3]; in other words, it is how important the magic trait actually is for speciation (or was, to be precise, since the definition is retrospective). The main point of our letter was to underline that this question of effect size is actually the important empirical question in need of investigation. "Magicness" may prove somewhat of a red herring, since, as described above, it's almost impossible to prove with certainty that a given trait is magic, and yet at the same time probably most or even all traits are in fact magic. We coined a term for the (perhaps very common) magic traits of very small effect: "squib traits." Like Kisdi's term "muggle trait," this comes from the Harry Potter world; a squib is a person who is born of magical parents, and who is magic in a non-useful sense (squibs can see Hogwarts, for example), but who cannot actually perform magic. A squib trait is like a Chevy on cinderblocks; it may be magic, but it ain't gonna take you anywhere (Figs. 4, 5).

Sadly, TREE nixed our coinage of "squib trait" (apparently there are still people somewhere who haven't read the Harry Potter books, and so the reference might be confusing :->), so we used the term "trivial magic trait" (versus "important magic trait") instead. Eva Kisdi's term "muggle trait" hasn't made it into the primary literature yet either. Bonus points to the first person who manages to get these terms into print! :->
I've done a lot of arguing about the definition of "magic trait" here, because I think it's interesting and it leads the discussion through many important issues, but it's the definition we've got, and it may be as a good a definition as any other that anybody could have come up with. What we really wanted to emphasize with our letter is: the definition is what it is, but the important questions in speciation do not revolve around the distinctions at the line between magic and non-magic. Rather, the important questions are things like:
Servedio et al., in their reply [5] to our letter, offer up some strong counterarguments to some of this, and remind the reader, in other cases, that they did give some consideration to these issues in their original paper (although not, we felt, enough consideration). But since this is my blog entry, and since it's already too long, I think I'll stop here, and let you, gentle reader, pursue the thread of argument further if you wish. And perhaps we can get a guest post from one of Servedio et al., to give you their own perspective!
Posted by: Benjamin C. Haller
References:
1. Schluter, D. (2001). Ecology and the origin of species. Trends in Ecology and Evolution 16, 372-380, doi:10.1016/S0169-5347(01)02198-X
2. Gavrilets, S. (2004). Fitness Landscapes and the Origin of Species. Princeton University Press
3. Servedio, M.R., Van Doorn, G.S., Kopp, M., Frame, A.M., Nosil, P. (2011). Magic traits in speciation: 'magic' but not rare? Trends in Ecology and Evolution 26, 389-397, doi:10.1016/j.tree.2011.04.005
4. Haller, B.C., De Léon, L.F., Rolshausen, G., Gotanda, K.M., Hendry, A.P. (in press). Magic traits: distinguishing the important from the trivial. Trends in Ecology and Evolution, doi:10.1016/j.tree.2011.09.005
5. Servedio, M.R., Van Doorn, G.S., Kopp, M., Frame, A.M., Nosil, P. (in press). Magic traits, pleiotropy and effect sizes: a response to Haller et al. Trends in Ecology and Evolution, doi:10.1016/j.tree.2011.09.011
Supplemental data:

Here's a specific empirical example: one of Darwin's finches, the Medium Ground Finch (Geospiza fortis) of the Galápagos (Fig. 1; discussed more in the previous post on this blog). We'll consider these birds as an example of "speciation in progress," but of course this means that it might not be an example of speciation at all – the early stages of divergence observed in this system may never "run to completion" and produce two fully distinct species. But because speciation is a process, it's best understood by looking at it as it's happening; by the time it's complete, much of the evidence regarding exactly how the process unfolded has vanished.

Figure 1. The Medium Ground Finch (Geospiza fortis), showing a
small-beaked individual (left) and a large-beaked individual (right).
Photo courtesy of De Léon, L.F.
To make a long story short, and glossing over many important details (see that previous blog entry and [1-4] for further discussion and cites), in the Galápagos there are small seeds and large seeds. The finches eat the seeds, and their beak size matters for this purpose because a large beak can generate more bite force than a small beak. So small-beaked birds are better suited to cracking small seeds, and large-beaked birds are better suited to cracking large seeds. The environment, by offering up these two seed types, is therefore applying divergent selection to the finches, and the finches, obediently following Darwin, have diverged adaptively: there are small-beaked morphs and large-beaked morphs, and the phenotypic distribution is bimodal, meaning there are relatively few medium-beaked morphs.
So that's great. Divergent selection, divergent adaptation – speciation? Not so fast. The problem is that sexual species, like these finches, mate, and when they mate, their offspring inherit chromosomes from both parents, which leads (in the typical case) to intermediate phenotypes. In other words, when a big-beaked finch mates with a small-beaked finch, their hybrid offspring will tend to be medium-beaked. So sexual reproduction pulls a population back together, opposing the diversifying force of divergent selection. This is the reason why the most commonly used definition of "species," the "biological species concept," defines "species" as being (mostly) reproductively isolated; if you're not reproductively isolated, then there's always the possibility that your incipient diversity will collapse back into a single phenotype due to mating.
How can this problem be overcome? This is the third and final step of ecological speciation, as I set it out at the beginning of the post: divergent adaptation must somehow lead to reproductive isolation. The finches seem to manage this: in some populations, there are relatively few medium-beaked morphs. There are lots of ways in which this can be achieved, and that's really what I want to talk about in this blog entry. Back in 2004, Sergey Gavrilets coined the term "magic trait" in a now-famous passage:
In the models considered so far in this chapter, speciation requires the establishment of correlations between the DS [divergent/disruptive selection] alleles and the NM [non-random mating] alleles. This requirement disappears if the same set of loci pleiotropically control both the trait subject to DS and the trait used in mating, or if the same trait underlies both processes... In this section, I consider several models, which I call magic trait models, describing such situations. [2, p. 368; emphasis in original]This passage actually refers to exactly the problem discussed above. In order to speciate, you need to diverge in the alleles that are under divergent selection – the "DS alleles." You also need to establish non-random mating somehow, so that the small-beaked guys don't mate with the big-beaked guys; in the models being discussed by Gavrilets, this is achieved by diverging in "NM alleles" that affect mate choice (for example, one NM allele might be expressed as a preference to mate with small-beaked finches, and another NM allele might be expressed as a preference to mate with large-beaked finches). Finally, for this to result in speciation, there must be the "establishment of correlations" between these two types of divergence: the non-random mating must be aligned with respect to the trait under divergent selection. It does you no good to diverge into groups that prefer small versus large beaks, if what the population is actually diverging in ecologically is red plumage versus blue plumage. Similarly, it does you no good if the individuals with small beaks end up with the allele coding for preference for large beaks, and vice versa. That's the problem in a nutshell: how to establish this correlation between DS alleles and NM alleles ("linkage disequilibrium" is the technical term).
In that passage, Gavrilets gave a solution as well: establishing that correlation is much easier if the loci under divergent selection and the loci that affect non-random mating are one and the same. Then divergent adaptation leads inevitably to non-random mating that allows the divergence to be maintained. It's like magic! (Fig. 2)

Figure 2. Magic, performed in this case by Harry Potter.
But there are several things about this definition that are hard to pin down, and so confusion reigned o'er the land until Servedio et al. bravely stepped into the fray [3]. In their much-needed paper, they clarify this definition in several important respects. First of all, there's the confusion over whether a magic trait is a phenotypic trait, as commonly imagined with reference to traits like beak size (and there are many other examples; see [3]), or whether it's actually a "magic gene" that governs such a trait, or perhaps even a set of "magic genes" (note Gavrilets' use of the word "loci," plural). I'll let you read [3] to see how they handled this hot potato. Second of all, there was a thorny problem regarding exactly which sorts of traits would be considered "magic" and which would not. Servedio et al. resolved this by defining two types of magic traits, "automatic" and "classic," which I'll explain with reference, again, to G. fortis.
Consider a large-beaked finch and a small-beaked finch that mate and have a medium-beaked baby finch (aww, isn't it cute?): that baby finch is the problem, and it must be eliminated. How to eliminate it? (I am speaking loosely for rhetorical purposes; I mean that it must be eliminated if divergence is to proceed, but of course G. fortis is not planning all this, nor is evolution guided somehow towards a goal of speciation.) One mechanism is via selection against that hybrid. If there are no medium-sized seeds in the system, then our medium-beaked bird will probably be outcompeted, will have low fitness, and will be relatively likely to die before reproducing. Through this mechanism, beak size is an "automatic magic trait"; reproductive isolation occurs, via selection against hybrids, as an automatic consequence of beak size being under divergent selection. There are other mechanisms by which magic traits can be automatic, involving spatial or temporal isolation, habitat choice, and some kinds of pollinator-related dynamics; see [3] for more details.
The other way that this undesirable baby bird can be eliminated (again, speaking loosely) is by not having it in the first place, through mate choice. If small-beaked finches prefer to mate with other small-beaked finches, and large-beaked finches prefer to mate with other large-beaked finches, then fewer hybrids will be produced (perhaps even none). And in fact, G. fortis does seem to mate assortatively, small with small and large with large, both because of a visual preference based on beak size, and because of imprinting on parental song, which, because beak size affects song, leads to a preference for mates with a beak size similar to one's own. This (both of these mechanisms of assortative mating based on beak size) makes beak size also a "classic" magic trait, by the definition of Servedio et al. The key distinction is that the non-random mating here is not inevitable or automatic; even with beak size under divergent selection, there is no a priori reason to think that that will lead to assortative mating. Instead, a separate genetic mechanism of some kind must be in place that establishes the rules that make it work, a mechanism that produces behaviors such as "I prefer to mate with males that have large beaks" or "I prefer to mate with males that sing a song like the one I heard my father sing when I was a baby." (I am here glossing over a further complication, between one-allele and two-allele mechanisms; see [3] and Felsenstein 1981). Once those rules, saying in one way or another "use beak size as a male mating cue," are set up (and they may have been set up in an ancestral species, long before G. fortis evolved), beak size acts as a magic trait; but since the use of beak size as a mating cue depends on that separate, external mechanism, it can be broken down, so it is not "automatic," it is "classic."
So far so good. Now our finches are 1) under divergent selection, 2) they exhibit adaptive divergence in response, and 3) they can preserve that adaptive divergence, to at least some extent, because the trait that is diverging, beak size, also "magically" produces assortative mating through both an automatic mechanism (selection against medium-beaked hybrids) and two classic mechanisms (female preference for particular beak sizes, and female preference for songs similar to the parental song they imprinted on). So... speciation? Again, not so fast.
The problem is that there are still various ambiguities and problems surrounding the "magic trait" definition, and so I and four co-authors wrote a letter to TREE [4] in response to Servedio et al. (and Servedio et al. replied [5]). Let's look at these difficulties, continuing with G. fortis as our example (see [4] for cites in support).
First of all, it's hard to decide whether beak size in G. fortis is truly "magic" or not, even by the clarified definition of Servedio et al. In G. fortis, divergent selection is only present in some years (and only at some sites); in other years (or at other sites), particularly when seeds are plentiful and so competition is low, medium-beaked individuals seem to do fine. So the black-and-white requirement of a "trait under divergent selection" is fulfilled only some of the time; is that good enough or not? In our letter, we propose that the "magicness" of magic traits is really the result of a trait-environment interaction. Beak size is not a "magic trait" unless it is placed in an environment that subjects it to divergent selection. In other environments, it is a "muggle trait," a trait which influences non-random mating but which is not presently under divergent selection; if a muggle trait is subjected to divergent selection, it becomes a magic trait. "Muggle trait" is a term coined by Eva Kisdi following the Harry Potter terminology in which "muggles" are people that are non-magical (Fig. 3). Similarly, such "magic environments" are only magic for organisms with a beak size trait that is used as a mating cue. So there will be, then, a spatiotemporal mosaic of "magicness" in those spots where the trait and the environment come together. It doesn't really make sense to talk about a "magic trait" except with reference to the "magic environment" it is in.

Figure 3. Muggles (unidentified); note their
aposematic coloration, perhaps Batesian mimicry.
aposematic coloration, perhaps Batesian mimicry.
Whether beak size in G. fortis is magic is unknown for other reasons as well. The "automatic" part of the non-random mating, since it depends upon selection against hybrids, occurs only for some years/sites, as just explained. The "classic" aspects of non-random mating in G. fortis depend upon how consistently, through time and across space, beak size is used as a mating cue, and that has not been studied well enough to draw firm conclusions, to my knowledge; it may be, for example, that beak size is only important for mate choice in lean years. The requirement of pleiotropy is also on slightly shaky ground, although it's better established than other aspects. Beak size certainly seems to be the trait under divergent selection, and seems also to be the trait used as a mating cue, and so pleiotropy seems guaranteed (since beak size and beak size are the very same phenotypic trait). But the hypothesis that beak size is the true trait subject to both divergent selection and assortative mate choice (rather than a correlated trait) can only really be shown conclusively with manipulative experiments (altering beak size somehow and observing the effect on fitness and mate choice) that are impossible to conduct since the Darwin's finches are protected species; so pleiotropy is not conclusively proved.
So we can't actually say whether beak size is magic or not; indeed, we can't even say with certainty whether any of the three definitional requirements are satisfied, despite this being one of the most extensively studied species on the planet. But there's a problem in the other direction as well: it might be a little too easy to claim that beak size is magic. You see, the definition of "magic trait" says nothing about the magnitude of any of its requirements. Even the tiniest amount of "divergent selection" greater than zero suffices. Even the tiniest tendency towards "non-random mating" above panmixia suffices. And even the tiniest amount of pleiotropy suffices, too – one could imagine an organism for which nose length is under divergent selection, toenail growth rate is used as a mating cue, and these two traits are largely governed by independent loci, but there is one locus, perhaps a regulatory gene of some sort, that has a tiny pleiotropic effect on both. That regulatory gene would then be a "magic trait" despite its very small pleiotropic contribution to the two phenotypic traits in question. So the definition draws a line, but at a place that isn't really related to the function of the thing being defined.
There are lots of cases where we work with such definitions. The car up on cinderblocks in your neighbor's yard had its wheels taken off a couple years back, and its engine is probably rusted firmly together; but it's still a "car," even though it isn't usable as transportation (Fig. 4). On the other hand, we often choose not to use such definitions. The "biological species concept" (BSC) definition of "species," set out by Ernst Mayr, originally required complete reproductive isolation between species, because otherwise two putative species might collapse back into a single species, in a case of the random mating problem we've been discussing, writ large. But in later writing Mayr (and the rest of biology) backed off from that requirement, as it became clear that many species in nature actually do hybridize with other species to a limited extent, and thus the absolute definition would not draw a line in a useful place. Rather than trying to draw a new, arbitrary line regarding just how much hybridization is allowable, the working definition of the biological species concept now essentially allows a vague "you know it when you see it" amount of hybridization. Despite this vagueness, it has proved more useful than the old, absolute definition. (Similarly, if you asked your neighbor "Hey, can I borrow your car?" and he gave you the keys to the Chevy on blocks, you would probably be unhappy – and if this happened frequently, you might start considering a different definition for the word "car"!)

Figure 4. A car.
Another reason the original BSC definition of "species" was loosened was that it wasn't testable. How do you prove that two species positively, absolutely, never, ever, interbreed? That's unprovable, and so the definition, used strictly, was useless. Similarly, how do you prove that a given trait 1) is not subject to even the tiniest smidge of divergent selection, 2) absolutely never affects mate choice, and 3) is not governed, even to the tiniest degree, by pleiotropy at any locus? You could never prove such a proposition, for any trait, and so for all we know, every single trait in every single (sexual) organism is magic. In fact, that's not even an unreasonable thing to assert. I think it might even be true. So maybe that's a problem.
This starts to get at the heart of the problem: effect size. The effect size of a magic trait is "how much the trait contributed to the evolution of increased reproductive isolation" [3]; in other words, it is how important the magic trait actually is for speciation (or was, to be precise, since the definition is retrospective). The main point of our letter was to underline that this question of effect size is actually the important empirical question in need of investigation. "Magicness" may prove somewhat of a red herring, since, as described above, it's almost impossible to prove with certainty that a given trait is magic, and yet at the same time probably most or even all traits are in fact magic. We coined a term for the (perhaps very common) magic traits of very small effect: "squib traits." Like Kisdi's term "muggle trait," this comes from the Harry Potter world; a squib is a person who is born of magical parents, and who is magic in a non-useful sense (squibs can see Hogwarts, for example), but who cannot actually perform magic. A squib trait is like a Chevy on cinderblocks; it may be magic, but it ain't gonna take you anywhere (Figs. 4, 5).

Figure 5. A squib, Argus Filch, with his cat, Mrs. Norris.
Sadly, TREE nixed our coinage of "squib trait" (apparently there are still people somewhere who haven't read the Harry Potter books, and so the reference might be confusing :->), so we used the term "trivial magic trait" (versus "important magic trait") instead. Eva Kisdi's term "muggle trait" hasn't made it into the primary literature yet either. Bonus points to the first person who manages to get these terms into print! :->
I've done a lot of arguing about the definition of "magic trait" here, because I think it's interesting and it leads the discussion through many important issues, but it's the definition we've got, and it may be as a good a definition as any other that anybody could have come up with. What we really wanted to emphasize with our letter is: the definition is what it is, but the important questions in speciation do not revolve around the distinctions at the line between magic and non-magic. Rather, the important questions are things like:
- How can we measure or estimate the expected effect size of traits in nature, given that the definition of "effect size" is retrospective?
- How common are magic traits of large effect size in nature, compared to traits that are either non-magic, or magic but of very small effect size (i.e. squibs)?
- How often is speciation driven by magic traits of large effect size, as compared to other mechanisms in which the magicness of traits is not important? (And, to follow up on a point in [5], are there plausible mechanisms in which magic traits of relatively small effect size are still important to speciation? But – if there are, then aren't those magic traits, by definition, of large effect size?)
- What drives the evolution of effect size itself; are there evolutionary mechanisms that tend to promote large effect sizes in magic traits, or were the magic traits of large effect size that we are beginning to find in some systems just produced by happenstance?
Servedio et al., in their reply [5] to our letter, offer up some strong counterarguments to some of this, and remind the reader, in other cases, that they did give some consideration to these issues in their original paper (although not, we felt, enough consideration). But since this is my blog entry, and since it's already too long, I think I'll stop here, and let you, gentle reader, pursue the thread of argument further if you wish. And perhaps we can get a guest post from one of Servedio et al., to give you their own perspective!
Posted by: Benjamin C. Haller
References:
1. Schluter, D. (2001). Ecology and the origin of species. Trends in Ecology and Evolution 16, 372-380, doi:10.1016/S0169-5347(01)02198-X
2. Gavrilets, S. (2004). Fitness Landscapes and the Origin of Species. Princeton University Press
3. Servedio, M.R., Van Doorn, G.S., Kopp, M., Frame, A.M., Nosil, P. (2011). Magic traits in speciation: 'magic' but not rare? Trends in Ecology and Evolution 26, 389-397, doi:10.1016/j.tree.2011.04.005
4. Haller, B.C., De Léon, L.F., Rolshausen, G., Gotanda, K.M., Hendry, A.P. (in press). Magic traits: distinguishing the important from the trivial. Trends in Ecology and Evolution, doi:10.1016/j.tree.2011.09.005
5. Servedio, M.R., Van Doorn, G.S., Kopp, M., Frame, A.M., Nosil, P. (in press). Magic traits, pleiotropy and effect sizes: a response to Haller et al. Trends in Ecology and Evolution, doi:10.1016/j.tree.2011.09.011
Supplemental data:

Figure S1. This is my nephew, who has a rather different
kind of magic trait action going on, as you can see. This
trait is hypothesized to be vulnerable to Kryptonite.
Happy Halloween, everybody!
kind of magic trait action going on, as you can see. This
trait is hypothesized to be vulnerable to Kryptonite.
Happy Halloween, everybody!
Wednesday, October 26, 2011
Homo sapiens influence micro-evolutionary processes in Darwin's finches
 Almost at the same time that the genus Homo was beginning to radiate in the African savanna, a single bird species from mainland South America was arriving to the Galápagos Islands. Millions of years later, Homo sapiens strive as the only living branch of the Homo radiation; in Galápagos, 14 bird species are the result of the finch radiation. Just as finches did in the past, the H. sapiens diaspora will soon manage to hit the Galápagos. Accidentally discovered by a lost Spanish bishop in 1535, the Galápagos have seen a multitude of visitors, ranging from pirates to whalers, explorers (Charles Darwin included) and prisoners in the 1700s and 1800s. By the 1950s, a few hundred people were living permanently in the Archipelago and the number of visitors was soaring. In the last few decades, these numbers have increased almost exponentially (Fig. 1). Although humans and finches are the products of independent evolutionary events, their encounter seemed inevitable. The consequences of such an encounter; however, were unimaginable.
Almost at the same time that the genus Homo was beginning to radiate in the African savanna, a single bird species from mainland South America was arriving to the Galápagos Islands. Millions of years later, Homo sapiens strive as the only living branch of the Homo radiation; in Galápagos, 14 bird species are the result of the finch radiation. Just as finches did in the past, the H. sapiens diaspora will soon manage to hit the Galápagos. Accidentally discovered by a lost Spanish bishop in 1535, the Galápagos have seen a multitude of visitors, ranging from pirates to whalers, explorers (Charles Darwin included) and prisoners in the 1700s and 1800s. By the 1950s, a few hundred people were living permanently in the Archipelago and the number of visitors was soaring. In the last few decades, these numbers have increased almost exponentially (Fig. 1). Although humans and finches are the products of independent evolutionary events, their encounter seemed inevitable. The consequences of such an encounter; however, were unimaginable.
 Figure 1. The growth of the H. Sapiens populations in the Galápagos Archipelago (from Watkins and Cruz 2007).
Figure 1. The growth of the H. Sapiens populations in the Galápagos Archipelago (from Watkins and Cruz 2007).
For instance, it is now believed that H. sapiens can affect the very evolutionary processes that promote adaptive radiation in Darwin's finches. This has recently been suggested for the Ground finches branch of this radiation. Specifically, our recent study published in Evolution suggests that human activities might be negatively impacting the initial stages of adaptive divergence within the medium ground finches (Geospiza fortis) on Santa Cruz Island (Fig. 2). Here is how the story goes.
Previous studies have shown that the medium ground finch (G. fortis), the morphologically most variable of the Darwin's finch species, shows evidence of morphological and genetic divergence on Santa Cruz Island. Specifically, this granivorous species shows a bimodal distribution in beak traits which seems to be the result of adaptive differences for food consumption, with a small beak morph feeding on small and soft seeds, and a large beak morph feeding on large and hard seeds. These two beak morphs also show genetic differences and are favoured by strong disruptive selection against birds with intermediate beak size. Although the origin of this divergence is not known, we do know that this bimodality varies spatially and temporally. For instance, at one site, El Garrapatero, the bimodality is strong but at other sites, such as Academy Bay and Borrero Bay, the bimodality is not evident. However, morphological data from the 60s and 70s suggest that the G. fortis population from Academy Bay was as bimodal in the past as El Garrapatero is in the present, but this bimodality has since weakened. Interestingly, this weakening in bimodality has occurred at the site with the highest H. sapiens population. Could H. sapiens be responsible for this loss of morphological divergence? And if so, what are the mechanisms by which this might be happening?


Figure 2. Small and large beak size morph of G. fortis from Santa Cruz Island (top panel). Beak size distribution in G. fortis from El Garrapatero (A), Academy Bay (B) and Borrero Bay (C) (photo credit: De León, L. F.; data from Hendry et al. 2006; De León et al. 2010).
The adaptive radiation in Ground finches supposes a tight link between morphological variation (beak size and shape), performance (bite force) and available resources (seed size and hardness). Therefore, any factor that can alter the underlying resource distribution could largely alter the adaptive landscape, promoting finch diversification. We argue that humans have drastically altered this adaptive landscape by introducing a large number of novel food resources (e.g., exotic plant species, potato chips, ice cream cones) which are readily exploitable by finch populations. We also argue that finch specialization on these novel resources could relax the association between morphology, ecology and performance, therefore weakening the selective forces responsible for the initial stages of divergence.
We tested these ideas by quantifying the diet of individual G. fortis, the availability of food resources and the associations between beak size, bite force, and seed size and hardness. The strength of these associations was then contrasted between the human perturbed (Academy Bay) and the unperturbed site (El Garrapatero). Both of the above predictions were supported by our data. Specifically, we found first, that finches at Academy Bay consistently exploit human-introduced food resources, and second, that the morphological-ecological-performance associations were generally weaker at the perturbed versus the unperturbed site. These results suggest that H. sapiens might indeed be eroding the bimodality in G. fortis on Santa Cruz Island (Fig. 3).


These results are important because we have pinpointed some of the causal mechanisms affecting the initial stages of divergence in one of the most celebrated examples of adaptive radiation. The same mechanisms could potentially be at work in other places in Galápagos and in other systems around the world. These results also call for new studies attempting to understand how H. sapiens can affect the micro-evolutionary processes which promote biodiversity.
It took millions of years for the finch and human diaspora to meet in Galápagos; however, only a few hundred years later, we can already see the effects of this encounter. What can we expect for the future? Although we don't have a definitive answer, we know that finches haven't stop radiating but neither has the wave of H. sapiens stopped arriving to the Galápagos (Fig. 4). If we don't pay attention to this interaction, we might realize that the evolutionary consequences of this encounter might not be that unimaginable after all.
 Figure 4. A new wave of H. sapiens visiting the Galápagos. Miss Ecuador, Señorita Claudia Schiess, arriving to Galápagos during field season 2011. She was followed by at least 100 more H. sapiens (me included). (photo credit: De León L. F.).
Figure 4. A new wave of H. sapiens visiting the Galápagos. Miss Ecuador, Señorita Claudia Schiess, arriving to Galápagos during field season 2011. She was followed by at least 100 more H. sapiens (me included). (photo credit: De León L. F.).
Posted by: Luis Fernando De León
Reference: De León, L. F., Raeymaekers, J. A M., Bermingham, E., Podos, J., Herrel, A., and A. P. Hendry. 2011. Exploring possible human influences on the evolution of Darwin’s finches. Evolution 65:2258-2272.
 Almost at the same time that the genus Homo was beginning to radiate in the African savanna, a single bird species from mainland South America was arriving to the Galápagos Islands. Millions of years later, Homo sapiens strive as the only living branch of the Homo radiation; in Galápagos, 14 bird species are the result of the finch radiation. Just as finches did in the past, the H. sapiens diaspora will soon manage to hit the Galápagos. Accidentally discovered by a lost Spanish bishop in 1535, the Galápagos have seen a multitude of visitors, ranging from pirates to whalers, explorers (Charles Darwin included) and prisoners in the 1700s and 1800s. By the 1950s, a few hundred people were living permanently in the Archipelago and the number of visitors was soaring. In the last few decades, these numbers have increased almost exponentially (Fig. 1). Although humans and finches are the products of independent evolutionary events, their encounter seemed inevitable. The consequences of such an encounter; however, were unimaginable.
Almost at the same time that the genus Homo was beginning to radiate in the African savanna, a single bird species from mainland South America was arriving to the Galápagos Islands. Millions of years later, Homo sapiens strive as the only living branch of the Homo radiation; in Galápagos, 14 bird species are the result of the finch radiation. Just as finches did in the past, the H. sapiens diaspora will soon manage to hit the Galápagos. Accidentally discovered by a lost Spanish bishop in 1535, the Galápagos have seen a multitude of visitors, ranging from pirates to whalers, explorers (Charles Darwin included) and prisoners in the 1700s and 1800s. By the 1950s, a few hundred people were living permanently in the Archipelago and the number of visitors was soaring. In the last few decades, these numbers have increased almost exponentially (Fig. 1). Although humans and finches are the products of independent evolutionary events, their encounter seemed inevitable. The consequences of such an encounter; however, were unimaginable.
For instance, it is now believed that H. sapiens can affect the very evolutionary processes that promote adaptive radiation in Darwin's finches. This has recently been suggested for the Ground finches branch of this radiation. Specifically, our recent study published in Evolution suggests that human activities might be negatively impacting the initial stages of adaptive divergence within the medium ground finches (Geospiza fortis) on Santa Cruz Island (Fig. 2). Here is how the story goes.
Previous studies have shown that the medium ground finch (G. fortis), the morphologically most variable of the Darwin's finch species, shows evidence of morphological and genetic divergence on Santa Cruz Island. Specifically, this granivorous species shows a bimodal distribution in beak traits which seems to be the result of adaptive differences for food consumption, with a small beak morph feeding on small and soft seeds, and a large beak morph feeding on large and hard seeds. These two beak morphs also show genetic differences and are favoured by strong disruptive selection against birds with intermediate beak size. Although the origin of this divergence is not known, we do know that this bimodality varies spatially and temporally. For instance, at one site, El Garrapatero, the bimodality is strong but at other sites, such as Academy Bay and Borrero Bay, the bimodality is not evident. However, morphological data from the 60s and 70s suggest that the G. fortis population from Academy Bay was as bimodal in the past as El Garrapatero is in the present, but this bimodality has since weakened. Interestingly, this weakening in bimodality has occurred at the site with the highest H. sapiens population. Could H. sapiens be responsible for this loss of morphological divergence? And if so, what are the mechanisms by which this might be happening?


Figure 2. Small and large beak size morph of G. fortis from Santa Cruz Island (top panel). Beak size distribution in G. fortis from El Garrapatero (A), Academy Bay (B) and Borrero Bay (C) (photo credit: De León, L. F.; data from Hendry et al. 2006; De León et al. 2010).
The adaptive radiation in Ground finches supposes a tight link between morphological variation (beak size and shape), performance (bite force) and available resources (seed size and hardness). Therefore, any factor that can alter the underlying resource distribution could largely alter the adaptive landscape, promoting finch diversification. We argue that humans have drastically altered this adaptive landscape by introducing a large number of novel food resources (e.g., exotic plant species, potato chips, ice cream cones) which are readily exploitable by finch populations. We also argue that finch specialization on these novel resources could relax the association between morphology, ecology and performance, therefore weakening the selective forces responsible for the initial stages of divergence.
We tested these ideas by quantifying the diet of individual G. fortis, the availability of food resources and the associations between beak size, bite force, and seed size and hardness. The strength of these associations was then contrasted between the human perturbed (Academy Bay) and the unperturbed site (El Garrapatero). Both of the above predictions were supported by our data. Specifically, we found first, that finches at Academy Bay consistently exploit human-introduced food resources, and second, that the morphological-ecological-performance associations were generally weaker at the perturbed versus the unperturbed site. These results suggest that H. sapiens might indeed be eroding the bimodality in G. fortis on Santa Cruz Island (Fig. 3).


Figure 3. A female G. fortis feeding on rice at Academy Bay (top panel). Correlation between morphology (beak size) and performance (bite force) at Academy Bay (bottom-left panel) and El Garrapatero (bottom-right panel) (photo credit: De León, L. F.).
These results are important because we have pinpointed some of the causal mechanisms affecting the initial stages of divergence in one of the most celebrated examples of adaptive radiation. The same mechanisms could potentially be at work in other places in Galápagos and in other systems around the world. These results also call for new studies attempting to understand how H. sapiens can affect the micro-evolutionary processes which promote biodiversity.
It took millions of years for the finch and human diaspora to meet in Galápagos; however, only a few hundred years later, we can already see the effects of this encounter. What can we expect for the future? Although we don't have a definitive answer, we know that finches haven't stop radiating but neither has the wave of H. sapiens stopped arriving to the Galápagos (Fig. 4). If we don't pay attention to this interaction, we might realize that the evolutionary consequences of this encounter might not be that unimaginable after all.
{kind=link}
 Figure 4. A new wave of H. sapiens visiting the Galápagos. Miss Ecuador, Señorita Claudia Schiess, arriving to Galápagos during field season 2011. She was followed by at least 100 more H. sapiens (me included). (photo credit: De León L. F.).
Figure 4. A new wave of H. sapiens visiting the Galápagos. Miss Ecuador, Señorita Claudia Schiess, arriving to Galápagos during field season 2011. She was followed by at least 100 more H. sapiens (me included). (photo credit: De León L. F.).Posted by: Luis Fernando De León
Reference: De León, L. F., Raeymaekers, J. A M., Bermingham, E., Podos, J., Herrel, A., and A. P. Hendry. 2011. Exploring possible human influences on the evolution of Darwin’s finches. Evolution 65:2258-2272.
Monday, October 17, 2011
Catastrophes and adaptations
Changes in the long-term mean state of climate variables (i.e., climate trends) and their consequences on evolutionary and adaptive processes in wild species have been intensively studied for more than 20 years. Climate change models show that the variance of climate variables like temperature or rainfall may change much more dramatically than their means and will thus intensify both the stochastic component of selection and extreme selective pressures. In the figure below, the dashed line represents the distribution of a climate variable (or phenotypic optimum, as often interpreted in eco-evo or quantitative genetics models) with climate change (solid line draws the distribution before climate change).

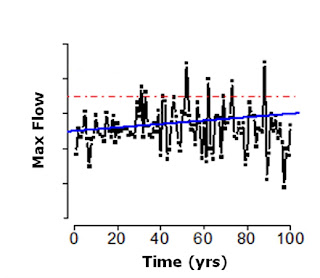

This intensification of weather extremes is emerging as one of the most important aspects of climate change, and the debate is expanding from an analysis of trends to an interest in extreme events such as periods of heavy rainfall or heat waves. There is compelling evidence that the frequency and severity of climate extremes have already increased in several regions. In the figure below, you can see a simple simulation showing the effects of an increase in the mean and variance of rainfall on the mean and variance of water flows in a hypothetical stream. The red line represents a 100-year flood (a flood with an expected recurrence interval of 100 years) pre-climate change (year 0), while the mean maximum flow over time is in blue. It is clear that with climate change we expect that what once was a 100-year flood will become, in fact, a 20-year flood.
In ecological models, how to define extreme events is a thorny question. A climate extreme could readily be represented by the tails of a distribution for a particular climate variable, but what constitutes a climate extreme will be highly dependent on the available climate record. An extreme climatic event (ECE) can be synthetically conceptualized to include “extremeness” in both the driver (climate) and the ecological response. In the ecological literature, ECE are often called catastrophes: that is, events characterized by statistical extremity, rapid onset, and abruptness relative to the life cycles of the organisms affected.
Many adaptations in life-history, morphological or behavioral traits may be associated with catastrophes, but the extent to which these extreme events have been shaping traits and might buffer natural populations against the increasing variance of climate variables is largely unknown. At this point we (ecologists) all know that evolutionary processes need to be incorporated into management programs in order to minimize biodiversity loss under rapid climate change, right?
My colleagues and I used marble trout as a model system to explore both the adaptive mechanisms helping population recovery after collapses and the role of catastrophes in shaping the life histories of affected populations. Marble trout is a resident salmonid endemic in Northern Italy, Switzerland and Slovenia, but only a few populations survive nowadays.

A conservation project started in 1993 in Slovenia thanks to the Biological Station of the Tour Du Valat, the Tolmin Angling Association, and a bunch of groups from different European universities. Ten pure populations have been discovered or created, and have been monitored with annuals sampling using a capture-recapture scheme. Marble trout live in harsh habitats which are frequently affected by punctuated catastrophes, such as storm-induced debris-flow and flash-floods, causing massive mortalities. Due to a debris-flow we lost the reintroduced population of Gorska in 2004.
Despite the absence of immigration, very low population sizes and genetic variability, these populations have persisted for centuries. This begs the question: what are the mechanisms allowing the persistence of populations affected by repeated catastrophes, and what are the consequences of repeated catastrophes in terms of evolution of life histories, genetic variability and phenotypic plasticity? According to theory, a range of traits can adaptively respond to recurrent catastrophes, such as: a) body growth rates, since they allow faster production after a population collapse, given the positive relationship between body size and fecundity; b) altered time of spawning; and c) morphological adaptations that can make fish avoid the floods or resist them, for example by staying close to the bottom of the stream bed using the pectoral fins. Moreover, density-dependent processes are also likely to shape life histories.
We focused our attention on body growth rates. However, the density-dependence of body growth and the large variations in population density often observed in stream-dwelling fish populations – even in the absence of catastrophic events – make the observation and disentanglement of the relative contributions of density and intrinsic body growth rates on the realized growth of individuals particularly challenging. We thus used an individual-based model to explore how density-dependent processes (survival and body growth) and different environmental conditions can create different selective environments for body growth rates. We investigated environments that are constant (with only demographic stochasticity), variable (including environmental stochasticity) and variable with the occurrence of unpredictable catastrophes (flash-flood events). In addition, we assumed that a negative (linear) relationship exists between survival rates and body growth. Model parameters were tuned for the marble trout population living in the stream Zakojska.
We found that in a variable environment with the occurrence of unpredictable catastrophes, there is selection for (relatively) fast growers, with a parallel maintenance of less variability in intrinsic body growth within the population. How does this selection for faster growers happen? Episodes of massive mortality trigger a chain of impacts leading to the selection of a limited range of intrinsic body growth rates in the affected population. Severe floods greatly reduce population size by flushing away or killing individuals, and the relaxation of density‑dependent pressure allows fish to rapidly increase their body size. In the years immediately following the episode of massive mortality, relatively fast growers can reach maturity earlier and, given the positive relationship between body length of females and fecundity, produce more eggs than slower growers. At the same time, the trade-off between body growth and mortality prevents the selection of very fast growers.
Our results showed that primarily flood events, not density-dependent processes, are driving the exclusion of slower growers from the population. The loss of slow growth traits through time could be considered to be local adaptation to the occurrence of high-amplitude stochastic events.
Among European countries, Slovenia has one of the highest frequencies of storms. Strong short-lived downpours or abundant several-day rainfalls can cause flooding, since the predominantly torrential character of watercourses means they rise very quickly. Data since the 1960s and preliminary explorations in our study area show a recent intensification, increased frequency and altered seasonality (autumn to spring) of catastrophic flooding events. The observed altered pattern is consistent with the predictions of the IPCC and of the European Commission’s Joint Research Centre (JCR). Evolution is usually thought to be a very slow process, something that happens over many generations, thanks to adaptive mutations. But environmental change due to anthropogenic causes such as climate change is happening very fast, and so adaptations, if they happen at all, may happen very fast.
To explore the effects of such rapid environmental change, we are now using an eco-genetic model to understand how the distribution of individual growth rates in a population of the freshwater salmonid marble trout evolves with an altered pattern of flood events, and whether the evolution of growth rates contributes to population persistence (evolutionary rescue). Stay tuned for our next results.
Vincenzi S, Crivelli AJ, Giske J, Satterthwaite WH, Mangel M in press. Selective consequences of catastrophes for growth rates in a stream-dwelling salmonid. Oecologia.
| |
Subscribe to:
Posts (Atom)
A 25-year quest for the Holy Grail of evolutionary biology
When I started my postdoc in 1998, I think it is safe to say that the Holy Grail (or maybe Rosetta Stone) for many evolutionary biologists w...

-
 As an editor, reviewer, supervisor, committee member, and colleague, I have read countless papers and proposals and have seen similarly co...
As an editor, reviewer, supervisor, committee member, and colleague, I have read countless papers and proposals and have seen similarly co... -
 Bloggy worms? Why worms? Let’s start this post with a small review. Since its start in 2009, the number of contributions on this blog ta...
Bloggy worms? Why worms? Let’s start this post with a small review. Since its start in 2009, the number of contributions on this blog ta... -
 By Dan Bolnick This past month, The American Naturalist published what I hope is the final step in the Editorial Board's evaluation of w...
By Dan Bolnick This past month, The American Naturalist published what I hope is the final step in the Editorial Board's evaluation of w...